
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по делопроизводству
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Рефераты по зарубежной литературе
Рефераты по логике
Рефераты по логистике
Рефераты по маркетингу
Рефераты по международному публичному праву
Рефераты по международному частному праву
Рефераты по международным отношениям
Рефераты по культуре и искусству
Рефераты по менеджменту
Рефераты по металлургии
Рефераты по муниципальному праву
Рефераты по налогообложению
Рефераты по оккультизму и уфологии
Рефераты по педагогике
Рефераты по политологии
Рефераты по праву
Биографии
Рефераты по предпринимательству
Рефераты по психологии
Рефераты по радиоэлектронике
Рефераты по риторике
Рефераты по социологии
Рефераты по статистике
Рефераты по страхованию
Рефераты по строительству
Рефераты по схемотехнике
Рефераты по таможенной системе
Сочинения по литературе и русскому языку
Рефераты по теории государства и права
Рефераты по теории организации
Рефераты по теплотехнике
Рефераты по технологии
Рефераты по товароведению
Рефераты по транспорту
Рефераты по трудовому праву
Рефераты по туризму
Рефераты по уголовному праву и процессу
Рефераты по управлению
Дипломная работа: Нейрофизиологические механизмы аномального сексуального поведения
Дипломная работа: Нейрофизиологические механизмы аномального сексуального поведения
А.В.Елисеев, А.А.Ткаченко, Т.В.Петина, Ю.Е.Куниковский
Сексуальные реакции и состояния, сексуальное влечение и поведение являются, возможно, и особыми, но все же частными случаями эмоциональных реакций и состояний, мотиваций и "мотивированного" поведения. Поэтому корректный анализ и интерпретация как литературных, так и собственных данных, имеющих отношение к указанной проблеме, практически невозможны вне такого, более широкого физиологического контекста. Исходя из этого, в первом разделе будет предпринята попытка синтезировать определенные концептуальные подходы к объяснению нейрофизиологических механизмов эмоционально-мотивационной сферы и приложить их к конкретным данным, полученным в настоящее время на стыке нейрофизиологии, психофизиологии, психологии, психиатрии и сексологии.
1. Церебральный базис сексуального поведения
1.1. Роль подкорковых структур лимбической системы в регуляции эмоциональных и мотивационных процессов
Паттерны поведения и животных, и человека формируются в филогенезе исходя из необходимости обеспечения как группового, так и индивидуального выживания, для которого гомеостаз является ключевым фактором, связанным с рецепцией жизненно важных параметров внутренней и внешней среды, регуляцией температуры тела, питанием, избеганием боли и т.п. Значение гипоталамо-гипофизарной системы в этих процессах трудно переоценить. Функции гипоталамуса, как известно, заключаются как в рецепции биологически значимых внутренних констант, так и в интеграции моторных, вегетативных и эндокринных компонентов поведения, которое организуется и регулируется при непосредственном участии структур переднего мозга (Шефер Д.Г., 1971; Вейн А.М., Соловьева А.Д., 1973; и др.). Гипоталамус обеспечивает перерастание биологических потребностей в биологические мотивации, которые активизируют мозг, вызывая перестройку деятельности лимбических структур и передних отделов неокортекса (Судаков К.В., 1971; и др.). Таким образом, гипоталамус участвует в регуляторных процессах и на самых ранних стадиях организации поведения, и на более поздних этапах, когда окончательно оформляется внешне реализуемый ответ (Симонов П.В., 1987; и др.).
С точки зрения эволюционного подхода, эмоциональный тон ощущений, выражающийся в чувстве дискомфорта или удовлетворенности и т.п., нужен для субъективного отражения успеха или неудачи отдельной особи в достижении ею целей, значимых для индивидуального и видового выживания. Эти "базисные" чувства и эмоции могут быть и относительно "элементарными", и достаточно сложными. Причем установлено, что аффективные реакции и даже определенные индивидуальные особенности поведения остаются сохранными при разрушении у животных неокортекса, в отличие от структур лимбической системы (Hara K., Myers R.E., 1973; Murphy M.R. et al., 1981; и др.). Интересно, что это, в определенной степени, относится и к сексуальному поведению у животных, причем преимущественно у самок, у которых деструкция неокортекса существенно не нарушала паттернов сексуального поведения (Blumer D., Walker A.E., 1975; и др.). Учитывая данные факты, высказывается точка зрения, что и у животных, и у человека как собственно продукция и субъективное переживание основных эмоций (в том числе и чувств сексуальной удовлетворенности или неудовлетворенности), так и развитие "сенсорно-аффективного" самосознания, определяется прежде всего деятельностью мезэнцефалических и лимбических подкорковых структур и, уже опосредованно, связанных с ними структур неокортекса (Doty R.W., 1989; Houdart R., 1994).
Рассматривая вопрос о структуре и функциях отдельных подкорковых структур лимбической системы, можно выделить три ее основных компонента: хабенуло-интерпедункулярный комплекс, гиппокампальную систему и миндалевидный комплекс.
Функции хабенуло-интерпедункулярного комплекса в настоящее время исследованы недостаточно, однако ряд данных позволяет высказать предположение, что из всех компонентов лимбической системы деятельность именно структур хабенуло-интерпедункулярного комплекса могла бы быть наиболее тесно связана с формированием собственно эмоций, как особых субъективных психических феноменов. В пользу этого предположения говорят следующие факты. Во-первых, связи данного комплекса с подкорковыми структурами (Jones E.G., 1985; Doty R.W., 1989; и др.), где осуществляется как рецепция тех или иных мотивационно и эмоционально-значимых гомеостатических показателей, так и перцепция "врожденных" (безусловно-рефлекторных) чувственных стимулов. Во-вторых, связи структур комплекса с нейрохимическими регуляторными системами (Azmitia E.C., Gannon P.J., 1986; Motohashi N. et al., 1986; и др.), что могло бы объяснить эффективность тех или иных фармакологических агентов при лечении аффективных расстройств и т.п.. В-третьих, полушарная асимметрия проекций комплекса (Mandell A.J., Knapp S., 1979; Doty R.W., 1983; и др.), что соотносится с полушарной "эмоциональной" асимметрией головного мозга. И, в-четвертых, данные о возникающих при стимуляции ряда структур этого комплекса субъективных эмоциональных переживаниях, в частности, чувстве эйфории и хорошего самочувствия (Schaltenbrand G. et al., 1973; и др.).
Гиппокампальная система, согласно данным многих авторов (Hirsh R., 1974; Виноградова О.С., 1975; Olton D.S. et al., 1979; Muller R.U. et al., 1987; и др.), ответственна прежде всего за регуляцию сохранения в долговременной памяти и извлечение из нее под влиянием мотивационного возбуждения следов ситуаций и событий, имеющих значение для организации целенаправленного поведения.
Миндалевидный комплекс связан афферентными и эфферентными волокнами как с различными подкорковыми структурами, так и с алло- и неокортикальными зонами (Russchen F.T., 1986; Price J.L., 1986; Amaral D.G., 1987; Акмаев И.Г., Калимуллина Л.Б., 1993; и др.). Причем важно отметить, что анатомические и функциональные взаимосвязи между правым и левым миндалевидными комплексами, также как и между правой и левой гиппокампальными системами, отсутствуют (Wada J.A. et al., 1981; Pandya D.N., Seltzer B., 1986; Rosene D.L, Van Hoersen G.W., 1987; Wilson C.L. et al., 1990, 1991). Касаясь функций миндалевидного комплекса, можно констатировать, что хотя миндалина и представляет из себя комплекс ядер, выполняющих различные задачи, но, в целом, среди них превалируют функции, связанные с организацией и регуляцией эмоциональных и мотивационных процессов, агрессивности и пр. (Clemente C.D., Chase M.H., 1973; Valenstein E.S, 1973; Halgren E. et al., 1978; Valzelli L., 1978; Чепурнов С.А., Чепурнова Н.Е., 1981; Gloor P. et al., 1982; Симонов П.В., 1987; и др.). Среди эмоционально-мотивационных функций миндалины чрезвычайно важное место занимают анализ и оценка эмоционально-ассоциированных сенсорных образов. На это указывает то, что активность единичных нейронов в миндалине у животных может строго селективно зависеть от эмоциональной и мотивационной значимости появляющихся в поле их зрения стимулов (Leonard C.M. et al., 1985; Ono T. et al., 1983; Kendrick K.M., Baldwin B.A., 1987). Современные исследования с использованием позитронно-эмиссионной томографии (ПЭТ) также дают основание предполагать, что передние структуры височной доли (миндалина, гиппокамп и височные области) ответственны прежде всего за "оценочную процедуру" (evaluation procedure), которая определяет оценку эмоциональной значимости экстрацептивных сенсорных стимулов (Reiman E., 1996).
Таким образом, миндалевидный комплекс может рассматриваться в качестве одной из главных структур эмоционально-мотивационной системы. Миндалина, во-первых, получает и "эмоционально" анализирует поступающую, в частности, из височно-теменной коры сенсорную информацию об окружающем мире, сопоставляет ее с мотивационным возбуждением из гипоталамуса, а также воздействует на гиппокамп, который регулирует сохранение в долговременной памяти и извлечение из нее данных, необходимых для текущего "эмоционального" анализа. Во-вторых, миндалевидный комплекс "взвешивает" конкурирующие эмоции, которые формируются при сопоставлении конкурирующих потребностей с перспективами их удовлетворения, и выделяет доминирующую мотивацию. И, в-третьих, принимая во внимание все эти источники, миндалина инициирует организацию адекватного имеющейся ситуации поведения в лобно-центральных регионах головного мозга и, посредством влияния на гипоталамус и вегетативную нервную систему, создает этому поведению соответствующее гормональное и нейровегетативное обеспечение (Симонов П.В., 1987; Doty R.W., 1989; и др.).
1.2. Роль подкорковых структур лимбической системы в регуляции сексуального поведения
Переходя собственно к подкорковым нейрофизиологическим механизмам сексуальных реакций и поведения и учитывая вышесказанное, можно попытаться следующим образом интегрировать накопленные за последние десятилетия данные по этому вопросу.
Организация проводящих путей, передающих сенсорную информацию, вызывающую сексуальное возбуждение, аналогична другим соматосенсорным системам. Репрезентация генитальной чувствительности связана с парацентральными регионами коры головного мозга. Афферентные проекции поступают и в гипоталамус, и в другие подкорковые образования. Из парацентральных областей нервные волокна следуют в срединные лимбические структуры височной доли, т.е. в миндалину, гиппокамп и прилегающие области лимбической коры (Blumer D., Walker A.E., 1975).
Эфферентные же волокна направляются, в частности, в гипоталамус, где, по всей видимости, находятся реципрокно-связанные между собой гормонально-зависимые центры, инициализирующие (преоптическая область) или тормозящие (область маммилярных тел) собственно копуляторные сексуальные действия (Lisk R., 1966, 1967; Имелинский К., 1986), которые реализуются при участии структур базальных ганглиев и моторно-двигательных зон коры. Поэтому изменение функций гипоталамических структур (и соответственно, гипоталамо-гипофизарно-гонадной системы) может играть важную роль в развитии нарушений сексуальности. Так, при поражении или дисфункции гипоталамических структур отмечаются как гипер-, или чаще гипосексуальность (Kolarsky A. et al. 1967; Taylor D., 1969; Ellison J.M., 1982; Хаит Н.З. 1993; и др.), так и девиантные формы сексуального поведения (Kula K., 1986; Темин П.А. и др., 1988; Шостакович Б.В. и соавт., 1990; и др.).
По-видимому, формирование оргазмической эмоциональной реакции на сексуальную стимуляцию осуществляется посредством вовлечения в процесс септальной области (MacLean P.D., 1957; Heath R.G., 1964; Смирнов В.М., 1976; и др.), деятельность которой модулируется соответствующими влияниями из гипоталамуса и зависит от уровня половых гормонов (Michael R.P., 1962; и др.). На существенную роль септальной области в механизмах инициации оргазмических переживаний указывает и ее тесная структурно-функциональная связь с хабенуло-интерпедункулярным комплексом (Jones E.G., 1985; Doty R.W., 1989; и др.), функции которого при формировании "базисных" эмоций обсуждались выше.
В свою очередь, деятельность миндалевидного комплекса, как вытекает из сказанного ранее, могла бы обеспечивать формирование сложных паттернов сексуального поведения. Во-первых, организуя комплексную эмоциональную и мотивационную оценку наличной ситуации и сенсорных стимулов, условно-рефлекторно связанных с сексуальной сферой, и формирование и извлечение из памяти, в кооперации с височными областями неокортекса и гиппокампом, соответствующих "сексуальных" ассоциаций. И, во-вторых, инициируя или тормозя (посредством влияния на лобно-центральные зоны коры, гипоталамус и базальные ганглии) "сексуально-ориентированное" поведение, в зависимости как от существующей ситуации, так и от наличия и выраженности других конкурирующих мотиваций (что в целом должно тормозить и ограничивать собственно "биологическую" сексуальность).
Такое понимание функции миндалевидного комплекса в механизмах сексуального поведения подтверждается, с одной стороны, давно известными фактами наличия при синдроме Клювера-Бюси (т.е. при билатеральной деструкции миндалевидного комплекса) выраженной гиперсексуальности, и, с другой стороны, наличием гипосексуальности у больных височной эпилепсией, с превалирующей у них гиперактивацией лимбических структур височной доли (Taylor D., 1969; Blumer D., 1970; Blumer D., Walker A.E., 1975; Темин П.А., Мухин К.Ю., 1989; и др.). В то же время более сложные дисфункциональные изменения в височных лимбических структурах (и прежде всего, по-видимому, в миндалине) при височной эпилепсии могут сопровождаться и различными сексуальными девиациями, такими как фетишизм, эксгибиционизм, вуайеризм и пр. (Kollarsky A. et al., 1967; Blumer D., Walker A.E., 1975; Ellison J.M., 1982; и др.).
Крайне велико значение лимбических структур и в механизмах сексуальной агрессии (которая у животных может являться компонентом сексуального поведения). Так, L. Valzelli (1978), рассмотрев результаты множества экспериментов на животных в этой области, пришел к заключению, что структуры гипоталамуса, вентрального гиппокампа и центромедиальной миндалины инициируют, а структуры септальной области, базолатеральной миндалины и цингулярной коры - тормозят проявления такого поведения. У человека повреждение лимбических структур височных долей также имеет значение в патофизиологических механизмах аномального агрессивного поведения (Garza-Trevino E.S., 1994; и др.).
Рассматривая роль подкорковых лимбических структур в организации сексуального поведения, можно отметить, что, судя по всему, именно на подкорковом уровне определяются основные, "базисные" различия как в паттернах сексуальной активности, так и в соответствующих соматических признаках и характеристиках между особями мужского и женского пола. Эти различия в паттернах сексуального поведения обусловлены половой дифференцировкой ряда лимбических структур, способных на ранних этапах онтогенеза запечатлевать детерминирующее влияние половых стероидов, и, в частности, некоторых ядер преоптической области, гипоталамуса и миндалевидного комплекса, которые контролируют секрецию половых гормонов и половое поведение (Акмаев И.Г., Калимуллина Л.Б., 1993; и др.)
Так, известно, что существуют половые различия в организации центров, регулирующих секрецию гонадотропинов. "Женский" центр локализован в преоптической области мозга (в медиальном преоптическом ядре) и переднегипоталамической области (в переднем гипоталамическом ядре), а "мужской" - в области аркуатного ядра (Barraclough C.A, Gorski R.A, 1961). Выявлены половые различия в размерах ядер преоптической области мозга (Gorski R.A. et al., 1978), некоторых ядер гипоталамуса и миндалевидного комплекса (Matsumoto A., Arai Y., 1983; Supprian T. et al. 1996; и др.). В частности, у особей мужского пола отмечаются достоверно большие размеры секдиморфного ядра преоптической области (Allen L.S. et al., 1989; и др.), медиальной задней области ядра ложа конечной полоски (Del Abril A. et al., 1987), ряда ядер кортикомедиальной группы миндалевидного комплекса (Mizukami S. et al., 1983; Акмаев И.Г., Калимуллина Л.Б., 1993), в то время как у особей женского пола выявлены значимо большие размеры некоторых ядер базолатеральной группы нейронов миндалины (Акмаев И.Г., Калимуллина Л.Б., 1993). Однако физиологическая значимость этих находок, как и соответствующие им нейрофизиологические механизмы, еще ждут своего прояснения.
1.3. Роль полушарных лимбико-кортикальных систем в регуляции эмоциональных и мотивационных процессов
Давно известно, что эмоциональные процессы существенно латерализованы, но мнения о характере такой латерализации разноречивы. Причем весь массив полученных результатов может быть условно разделен на две части.
Во-первых, ясно, что функции правого полушария доминантны как в отношении перцепции и когнитивного анализа "невербальной" личностно- и эмоционально-ассоциированной сенсорной информации, так и в отношении организации и регуляции эмоциональной экспрессии, причем независимо от знака соответствующей эмоции. В частности, на это указывают данные психофизиологических экспериментов, продемонстрировавших как лучшее восприятие и когнитивный анализ в правом полушарии эмоциональной мимики (Magnussen S. et al. 1994; Lane R.D. et al. 1995; и др.) и эмоциональной окраски голоса (Bryden M.P. et al., 1991; и др.), так и правополушарную доминантность в регуляции мимической экспрессии (Moreno C.R. et al., 1990; Christman S.D., Hackworth M.D., 1993). Об этом же говорят и результаты наблюдений за больными с поражением правого полушария, у которых были больше нарушены процессы опознания эмоциональной мимики (Mandal M.K. et al., 1991; Borod J.C. et al., 1993) и прозодических компонентов речи (Lalande S. et al. 1992; Ackermann H. et al. 1993; Starkstein S.E. et al. 1994), а также мимическая (Blonder L.X. et al., 1993) и вербальная (Bloom R.L. et al., 1992) эмоциональная экспрессия. Кроме этого, установлено, что правое полушарие в большей степени ответственно за опознание персональных характеристик лиц, голосов и т.п. (Van Lancker D., 1991 и др.).
Во-вторых, имеется большой массив данных, указывающих на то, что правое и левое полушария связаны с эмоциональными процессами различного знака.
Так, правое полушарие доминантно при перцепции и когнитивном анализе отрицательной эмоционально-ассоциированной сенсорной информации. Это показано в отношении отрицательной эмоциональной мимики (Szelag E., Fersten E., 1991; Johnsen B.H., Hugdahl K., 1993) и визуальных отрицательных эмоциональных стимулов (Wittling W., Roschmann R., 1993; и др.). Соответственно, повреждение правого полушария больше нарушает оценку отрицательной эмоциональной мимики (Mandal M.K. et al., 1991). Активация правого полушария усиливает отрицательную эмоциональную окраску голоса (Schiff B.B., Lamon M., 1994), а его инактивация сопровождается положительными эмоциями (Lee G.P. et al. 1993; и др.).
В то же время установлено, что левое полушарие, напротив, лучше оценивает положительную эмоциональную мимику (Bryson S.E. et al., 1991; McDowell C.L. et al., 1994). Кроме этого, левое полушарие более активировано при оценке положительной эмоциональной мимики (Gur R.C. et al., 1994), при восприятии приятных визуальных стимулов (Jones N.A., Fox N.A., 1992), при индуцированных положительных эмоциях (Kochi K. et al., 1996). Соответственно, инактивация левого полушария сопровождается отрицательными эмоциями (Lee G.P. et al. 1993; Christianson S.A. et al., 1993 и др.).
Таким образом, можно резюмировать, что согласно одним данным, правое полушарие в большей степени связано с перцепцией, когнитивным анализом и выражением эмоциональной экспрессии, независимо от знака эмоции. Согласно же другим данным, правое полушарие связано с формированием преимущественно отрицательных эмоций, а левое - положительных эмоций.
Объяснение этого противоречия могло бы, в частности, исходить из того, что преимущественная переработка эмоционально-ассоциированной сенсорной информации (мимической, прозодической и т.п.) и регуляция эмоциональной экспрессии в правом полушарии обусловлены не столько тем, что данные сенсорные или моторно-двигательные паттерны связаны именно с эмоциями, сколько тем, что правое полушарие преимущественно специализировано как раз на тех способах обработки информации, с помощью которых в основном анализируется и организуется эмоциональная экспрессия. Однако вопрос о том, в чем заключаются принципиальные различия этих способов переработки информации в левом и правом полушариях головного мозга, пока еще не ясен. Как указывают С.Спрингер и Д.Дейч (1983), современная нейрофизиология хотя и дает некоторые ответы на вопрос о том, что может, а чего не может делать каждое полушарие, но все еще далека от понимания причин и механизмов специализации полушарий. Представляется, что в контексте данной работы наиболее адекватными и перспективными были бы следующие подходы. Это, во-первых, представления о том, что в левом полушарии более развиты нейрофункциональные системы, обеспечивающие перцепцию и когнитивный анализ "типичного", "общего", "абстрактного" в сенсорных образах, а в правом полушарии - "комплементарные" процессы выделения и когнитивного анализа "особенного", "индивидуального" и "конкретного". Во-вторых, это теоретические положения о большей развитости в левом полушарии нейрофизиологических систем, обеспечивающих формирование "обычных", "типичных", а в правом полушарии - "особых", "индивидуальных" поведенческих паттернов.
Детальное и всестороннее обсуждение данной гипотезы выходит за рамки данной работы. Однако можно отметить, что при этом получают свое объяснение некоторые вопросы, связанные с механизмами формирования у человека в филогенезе и онтогенезе доминантности левого или правого полушарий в отношении ряда аспектов психической деятельности. В частности, процессов обработки вербальной или невербальной информации, рационального или интуитивного мышления, сознательного или бессознательного, эмоций, речи, особенностей личности и т.д. (Спрингер С., Дейч Г., 1983; Брагина Н.Н., Доброхотова Т.А., 1988; Зенков Л.Р., 1994; и др.). Так, возможно, становится понятным формирование у человека специализации левого полушария одновременно и на анализе "вербального" (т.е. определенных абстрактных символов), "нейтрального" (т.е. обычного, стандартного экспрессивного паттерна), "социального" и "рационального" (т.е., поведенчески или когнитивно-стереотипного), и на организации в левом полушарии речи и рационализированного социально-стереотипного ("нормального") поведения. И, соответственно, получает свое объяснение формирование правополушарной доминантности как при анализе "невербального" (т.е. особого, конкретного в воспринимаемых сенсорных образах), "эмоционального" (т.е экспрессивного паттерна, отклоняющегося от обычного), "индивидуального" (нетипичного), и "иррационального" (нестереотипного), так и при формировании невербальной экспрессии и организации "особых", "эмоциональных" или "индивидуальных" (в том числе и "аномальных") поведенческих паттернов.
Что касается данных о связи левого полушария непосредственно с "продукцией" положительных, а правого полушария - отрицательных эмоций, то, как считает П.В. Симонов (1987), было бы упрощением думать, что "центры" положительных эмоций локализованы в левом полушарии, а "центры" отрицательных эмоций - в правом. Однако игнорирование огромного массива сведений о связи полушарий с эмоциями разного знака невозможно, а представления только об "информационном" генезе латерализованных эмоциональных эффектов плохо согласуются с современными нейроанатомическими и нейрофизиологическими данными о наличии тесных взаимосвязей между корковыми и лимбическими структурами. Поэтому наиболее адекватным и взвешенным представляется следующий подход. Так, если собственно процессы формирования эмоций, как указывалось ранее, являются функцией срединных подкорковых структур лимбической системы, то полушария, возможно, ответственны прежде всего за восприятие и когнитивный анализ эмоционально-ассоциированных сенсорных образов. В таком случае деятельность полушарных лимбико-кортикальных систем, разделенных на лимбическом (миндалина, гиппокамп) и связанных на корковом уровне, обеспечивает восприятие и анализ позитивно-ассоциированных сенсорных стимулов преимущественно в левом полушарии (что может опосредованно инициировать положительные эмоции), а анализ негативно-ассоциированных сенсорных образов - в правом (что, соответственно, может приводить к инициации отрицательных эмоциональных реакций и состояний).
Помимо связи собственно эмоциональных процессов с правым и левым полушариями необходимо, как представляется, остановиться и на возможной связи полушарий с "мотивационными" процессами или с тем, что в теории функциональных систем соотносится со стадией принятия решений (Анохин П.К., 1975; Судаков К.В., 1976), т.е. с процессами, благодаря которым на основе имеющихся потребностей, когнитивного анализа, эмоциональных оценок наличной ситуации и возможностей организации деятельности (т.е. таких компонентов предшествующей стадии афферентного синтеза, как мотивация, накопленный опыт, обстановочная сигнализация и пусковые стимулы) в лобных зонах коры головного мозга формируются адекватные поведенческие паттерны, или иначе, определенная "линия поведения" (Хомская Е.Д., 1987; Урываев Ю.В., 1996 и др.). Данные об этом аспекте функционирования эмоционально-мотивационных систем пока немногочисленны, однако чрезвычайно интересны. Так, в работе С.В. Мадорского (1982) выявлено, что поражение правого миндалевидного комплекса сопровождается значительным усилением потребности в достижении успеха, а поражение левого - возрастанием стремления к избеганию неудач. В другом исследовании обнаружено, что у лиц с "левополушарным" типом нейрофизиологического реагирования наблюдается чрезмерная потребность в поисковой активности (Аршавский В.В., 1990). А в работах J.H. Gruzelier и его сотрудников (Gruzelier J.H., 1994; Gruzelier J.H., Doig, A., 1996; и др.) показано, что так называемый "Активный" ("Active") синдром шизофрении (или фактор шизотипии), характеризуемый, в частности, повышенной поведенческой активностью, связан с относительно большей активацией левого полушария, а синдром (или фактор) "Избегания" ("Withdrawn") - с большей активацией правого полушария.
В свете этих данных вполне естественным выглядит предположение о том, что в левом полушарии должны доминировать лимбико-кортикальные системы, обеспечивающие "мотивационные" процессы организации "достижения" эмоционально-позитивного, а в правом полушарии, наоборот, - комплементарные нейрональные системы организации "избегания" эмоционально-отрицательного. Подобных взглядов на наличие и функции латерализованных "мотивационных" систем в головном мозге придерживается в настоящее время и ряд ведущих нейро- и психофизиологов. Так, в модели J.T. Cacioppo (1996) предполагается, что позитивный и негативный мотивационные процессы, определяющие соответствующие аффекты и эмоции, различны (стохастически и функционально независимы). R.J. Davidson (1992) основную роль в обеспечении поведения, ориентированного на "достижение" (approach) и "избегание" (withdrawal), приписывает, соответственно, левым и правым лобным и передним височным регионам полушарий головного мозга. В концепции D.M. Tucker et al. (1995) указывается на факты, свидетельствующие о существовании двух лимбико-фронтальных систем, - дорзальной и вентральной, - имеющих различную структуру и происхождение и асимметрично представленных в головном мозге (соответственно больше в левом и правом полушариях). Предполагается, что дорзальные лимбико-фронтальные системы преимущественно задействованы в гедонистической оценке, в формировании социальных взаимосвязей, в холистическом и импульсивном типе контроля за поведением, а вентральные лимбические проводящие пути от миндалины к орбитальной фронтальной коре в большей степени вовлечены в осуществление "тормозного", "ограничивающего" типа поведения.
1.4. Половой диморфизм в организации полушарных лимбико-кортикальных систем регуляции поведения
Данные о половом диморфизме в организации и функциях полушарных корковых структур мозга в норме скудны и противоречивы. При их рассмотрении создается ощущение, что различия в полушарной асимметрии между мужчинами и женщинами имеют достаточно тонкий характер.
Так, в частности, сведения об относительно больших размерах задней части мозолистого тела у женщин (de Lacoste-Utamsing M.-C., Holloway R.L., 1982; и др.), которые, впрочем, подтверждаются не во всех исследованиях (Supprian T. et al., 1996), могут указывать на то, что при решении тех или иных задач женщины используют оба полушария в большей степени чем мужчины, что могло бы объяснить относительно меньшую выраженность у них как "лево-", так и "правополушарных" нарушений при локальных латерализованных поражениях головного мозга и сравнительно большую билатеральность в организации функционирования их мозга (McGlone J., 1977, 1980; Bryden M.P., 1982; и др.).
Кроме этого, различия в церебральной организации между мужчинами и женщинами неодинаковы в разных областях мозга, т.е. имеет место половой нейрональный мозаицизм - neural sexual mosaicism (Witleson S.F., 1991). Отсюда вытекает то, что эти различия могут быть обусловлены и связанными с полом особенностями функционирования разных, и прежде всего передних и задних, регионов. Так, показано, что у мужчин значительно чаще встречаются выраженные афазии и апраксии при поражении задних, а у женщин, наоборот, - передних отделов левого полушария (Kimura D., Harshman R.A., 1984). Если исходить из того, что относительно большее нарушение функций должно наблюдаться при локальном левополушарном повреждении в большей степени левополушарно-латерализованной мозговой системы (так как возможностей для функциональной компенсации дефекта за счет структур сохранного контралатерального правого полушария в такой системе существенно меньше), то данные факты могут свидетельствовать об относительно большей латерализованности у мужчин задних (височно-теменных), а у женщин - передних (лобно-центральных) корковых систем регуляции речи и произвольных действий.
Соответственно можно предположить сравнительно большую развитость в обоих полушариях (и меньшую латерализованность) у мужчин лобно-центральных (связанных преимущественно с корковым уровнем регуляции моторно-двигательных процессов), а у женщин - височно-теменных нейрональных систем (отвечающих в основном за когнитивный анализ сенсорной информации). Последнее, по всей видимости, соотносится как с упоминавшимися выше сведениями о большей развитости у женщин именно задней части мозолистого тела (de Lacoste-Utamsing M.-C., Holloway R.L., 1982 и др.), так и с данными о меньшей анатомической асимметрии у женщин в заднем височном регионе коры головного мозга (Wada J.A. et al., 1975). В пользу данного предположения говорит и характер тех заданий, которые лучше выполняют мужчины или женщины (Кимура Д., 1992; и др.). Так, в частности, мужчины лучше выполняют задачи, требующие умственных манипуляций с предметами, математических рассуждений или некоторых точных двигательных навыков, т.е. способностей, тесно связанных с деятельностью как лево-, так и правополушарных лобно-центральных областей, женщины же обычно превосходят мужчин в быстроте визуальной идентификации сходных предметов, в арифметическом счете и запоминании ориентиров местности, т.е., в том, что преимущественно связано с функционированием височно-теменных зон и левого, и правого полушарий головного мозга (Хомская Е.Д., 1987; и др.).
Следуя этой же логике в отношении полового диморфизма в организации лимбико-кортикальных систем, регулирующих эмоциональные и мотивационные процессы, можно предложить следующую гипотезу. Согласно ей, половые различия в строении и функциях этих систем связаны с неодинаковой развитостью в обоих полушариях (и различной степенью латерализованности) у мужчин и женщин височно-теменных и лобно-центральных (соответственно "эмоциональных" и "мотивационных") компонентов данных систем.
Так, предполагается, что в норме у женщин менее латерализованы и более развиты в обоих полушариях височнотеменные лимбико-кортикальные системы, анализирующие и оценивающие сенсорные эмоционально-ассоциированные (и положительные, и отрицательные) стимулы. Причем вследствии того, что эти нейрональные системы более интенсивно развиваются в период раннего онтогенеза, они, по всей видимости, и более уязвимы при действии различных факторов, нарушающих процессы формирования структур головного мозга в этот период жизнедеятельности.
У мужчин же, возможно, менее латерализованы и более развиты в обоих полушариях лобно-центральные лимбико-кортикальные системы, определяющие собственно "мотивационные" процессы организации поведения (ориентированного и на "достижение", и на "избегание"). Эти лобно-центральные нейрональные системы у мужчин являются, вероятно, и более уязвимыми в период раннего онтогенеза, что должно проявляться в большей выраженности у лиц мужского пола дизонтогенетических нарушений данных систем, в частности, в виде формирования аномальных паттернов полушарной асимметрии в рассматриваемых областях головного мозга.
Рамки данной работы не позволяет здесь подробно рассмотреть всю аргументацию за и против данной точки зрения. Однако можно отметить, что в пользу этой гипотезы говорит ряд хорошо известных фактов, касающихся полового диморфизма эмоциональных, мотивационных и личностных характеристик и нарушений у человека. Так, во-первых, женщины на психологическом уровне обычно более эмоциональны и более ситуационно- и социально-детерминированы в своем поведении, в то время как мужчины, напротив, менее эмоционально чувствительны и экспрессивны и более "самостоятельны" и активны в выборе и реализации индивидуальных форм своего поведения, что сопровождается и различной иерархией ценностей у женщин и мужчин в отношении "чувства" и "дела" и пр. (Eysenck H.J., 1976; Кон И.С., 1989; Русалов В.М., 1993; Feingold A., 1994; и др.). И, во-вторых, как и должно быть в соответствии с данной гипотезой, у женщин существенно чаще встречаются различные дизонтогенетически обусловленные нарушения в аффективной сфере, в частности, биполярные расстройства (Ротштейн В.Г и др., 1997 и др.), в то время как расстройства влечений, в том числе и сексуального, отмечаются в основном у мужчин (Flor-Henry P., 1987; и др.).
1.5. Роль полушарных лимбико-кортикальных систем в регуляции сексуального поведения
Нейрофизиологическая концепция P. Flor-Henry (1987), касающаяся роли полушарных отношений в организации и регуляции сексуального поведения (в том числе и аномального), исходит из того, что полушарные нейрональные структуры непосредственно и достаточно жестко связаны с организацией тех или иных паттернов сексуальной активности. В этой концепции предполагается, что в левом полушарии формируются вербально-идеаторные схемы сексуального поведения, которые затем передаются в правое полушарие, где эти схемы преобразуются в соответствующие моторно-двигательные паттерны сексуальной активности. При этом возникновение аномальных форм сексуального поведения рассматривается как следствие дефицитарности нейрофизиологических процессов формирования в левом полушарии "нормальных" вербально-идеаторных схем сексуальной активности либо как нарушение передачи этих "моделей" в правое полушарие или как недостаточность правополушарных нейрофизиологических механизмов, воплощающих эти схемы в конкретные "сексуальные" действия.
Однако связь именно левого полушария с формированием "схем" сексуальной активности , как и то, что эти схемы обязательно должны быть "вербально-идеаторными", вызывает большое сомнение и противоречит данным многочисленных исследований механизмов сексуального поведения животных и человека, указывающих на то, что формирование паттернов сексуальной активности ("конкретно-чувственных" по своей природе) обусловлено прежде всего деятельностью подкорковых и лимбических структур головного мозга (и преимущественно право-, а не левополушарных). Кроме этого, в концепции P. Flor-Henry (1987) полушарные механизмы сексуального поведения фактически рассматриваются изолированно, в отрыве от анализа огромного массива данных, указывающих на связь полушарных отношений с общими процессами регуляции эмоций и "мотивированного" поведения (разновидностью которых и являются "сексуальные" эмоциональные реакции и сексуальное поведение).
Поэтому более продуктивным и обоснованным мог бы быть иной концептуальный подход, который и будет использоваться нами в дальнейшем для интерпретации собственных и литературных данных по изучаемому вопросу. Согласно этому подходу полушарные нейрофизиологические механизмы регуляции сексуального поведения будут рассмотрены в контексте изложенных ранее современных представлений о структуре и функциях полушарных лимбико-кортикальных систем регуляции эмоциональных и мотивационных процессов. Это позволит в этой части главы достаточно логично структурировать уже имеющиеся в литературе данные и высказать ряд предположений, а в следующей части - интерпретировать результаты собственных исследований и сформулировать новую гипотезу психо- и нейрофизиологических механизмов формирования аномального сексуального поведения.
Во-первых, можно ожидать, что экспрессивное выражение сексуального возбуждения и оргазмическая реакция как особые (во многом непроизвольные) формы эмоциональной экспрессии должны быть связаны в большей степени с активацией правого полушария. И это находит свое подтверждение. Так, при оргазмической эпилепсии в подавляющем числе случаев очаг находится в правом полушарии (Flor-Henry P., 1983). При оргазме наблюдаются выраженные изменения ЭЭГ также преимущественно в правой париетальной области (Cohen H.D. et al., 1976). Эта же область гиперактивирована и при эротической стимуляции и переживании сексуального возбуждения (Tucker D.M., Dawson S.L., 1984).
Во-вторых, из предыдущего анализа вытекает, что левое полушарие могло бы быть связано преимущественно с интериоризацией и реализацией "нормальных" социальных стереотипов сексуального поведения, а правое полушарие - с усвоением и организацией "индивидуальных" (особых) форм и способов сексуальной активности, могущих в ряде случаев становиться и "аномальными". Следовательно, вполне логично ожидать в таком случае у лиц с отклонениями от социальных стереотипов сексуального поведения (с сексуальными девиациями) гиперактивации правого и/или гипоактивации левого полушария. Что в отношении гипоактивации левого полушария и нашло свое подтверждение в ЭЭГ-исследованиях лиц с эксгибиционизмом и педофилией, проведенных P. Flor-Henry с сотр. (1988, 1991). Вопрос о существовании правополушарной гиперактивации у лиц с аномальным сексуальным поведением оставался до последнего времени неясным, хотя в литературе имелись свидетельства наличия структурных нарушений в правом полушарии у сексуальных садистов (Langevin R. et al., 1988; Ether R. et al., 1996).
В-третьих, можно предположить, что анализ эмоционально-положительных ассоциативных "сексуальных" и "социальных" образов (символов жизненного и сексуального "успеха", эротики и пр.) должен осуществляться в большей степени в левом полушарии, а анализ аналогичных отрицательных ассоциативных образов (символов сексуальной неудовлетворенности и ущербности, импотенции и пр.) - в правом. Поэтому указанное выше изменение полушарных отношений в сторону сравнительно большей правополушарной активированности у лиц с парафилиями и должно, согласно данной модели, сопровождаться аффективными расстройствами субдепрессивного характера, дисфориями и т.п. И действительно, было показано, что парафилии нередко сочетаются с дистимическими, дисфорическими и генерализованными тревожными расстройствами (Coleman E., 1992; и др.).
В-четвертых, можно ожидать, что "мотивационные" процессы организации "достижения" положительных, в том числе и сексуально-ассоциированных, ощущений доминируют в левом полушарии, а процессы организации "избегания" неприятных социальных и сексуальных ощущений (чувств "социальной" и сексуальной неудовлетворенности или неполноценности) - в правом.
Комплексное нейрофизиологическое исследование лиц с аномальным сексуальным поведением, результаты которого будут изложены далее, позволило подтвердить данные предположения, также как и прояснить некоторые вопросы, связанные с патогенетическими и патофизиологическими механизмами парафилий.
2. Компьютеризированное ЭЭГ-обследование лиц с парафилиями
2.1. Нейрофизиологические паттерны полушарных, зональных и корково-подкорковых отношений у лиц с парафилиями
Обследовались лица мужского пола, находившиеся на стационарной судебно-психиатрической экспертизе в лаборатории судебной сексологии в ГНЦ СиСП им. В.П. Сербского в период 1993-1996 гг. после их привлечения к уголовной ответственности за сексуальные правонарушения. В результате клинико-психопатологического и сексологического исследования испытуемые были разделены на две группы.
Первую клиническую группу составили лица с парафилиями (ЛСП), диагноз которых ставился в соответствии с критериями DSM-III-R и МКБ-10. В основном среди форм парафилий отмечались садизм, педофилия и эксгибиционизм. Нередко встречались множественные расстройства сексуального предпочтения, при которых указанные виды парафилий сочетались с фетишистскими, вуайеристскими, мазохистическими тенденциями.
Вторую группу составили лица с аномальным сексуальным поведением без парафилий (ЛБП).
ЭЭГ-обследование сопоставимых по возрасту праворуких испытуемых мужского пола с аномальным сексуальным поведением (37 лиц с парафилиями и 43 - без парафилий) и группы нормы (ГН), состоящей из 49 праворуких психически и неврологически здоровых добровольцев мужского пола, производилось с учетом современных методических требований (Pivik R.T. et al., 1993) на компьютерном картографе MEDICID-3M (Куба) в 16 отведениях униполярно (с референтными закороченными ушными электродами) в четырех функциональных состояниях: при закрытых глазах (ЗГ), при открытых глазах (ОГ), при арифметическом счете в уме (СЧЕТ), т.е. при последовательном вычитании с закрытыми глазами, а также при выполнении зрительно-пространственного задания (ЗПЗ), т.е. при тесте Тейлора. В последующем с использованием стандартного программного обеспечения (DE-MEDICID) проводился спектрально-когерентный анализ полученных данных с вычислением в четырех диапазонах ЭЭГ (дельта, тета, альфа и бета-1) значений таких параметров, как логарифм абсолютной спектральной мощности (ЛАСМ) и межполушарная когерентность (МКОГ). Статистическая обработка (SPSS for Windows) предусматривала попарно групповое сравнение параметров ЛАСМ и МКОГ методом дисперсионного анализа (ANOVA), и в том случае, если выявлялось достоверное (p<0.05) влияние "группового" фактора, сравнение групп по частным средним (LSD-тест).
При анализе МКОГ и ЛАСМ у лиц с парафилиями был выявлен ряд характерных зональных и полушарных особенностей ЭЭГ.
Так, если сниженный (по сравнению с нормой) уровень межполушарной когерентности наблюдался у обеих клинических групп (у ЛСП - преимущественно в лобных и центральных зонах, а у ЛБП - больше в затылочных областях), то достоверное (p<0.05) повышение межполушарной когерентности в задневисочных и париетальных областях неокортекса при различных функциональных состояниях во всех ЭЭГ-диапазонах отмечалось только у лиц с парафилиями и по отношению к ГН, и по отношению к ЛБП (рис. 6.1). Учитывая, что для подкорковых структур таламо-лимбико-ретикулярного комплекса характерны преимущественно билатерально-синхронные влияния на кору головного мозга, с определенной степенью уверенности можно рассматривать увеличение показателя межполушарной когерентности в височных и париетальных областях в контексте изменения не только корково-корковых, но и корково-подкорковых отношений. Соответственно можно говорить о снижении у лиц с парафилиями степени относительной автономности деятельности височных (и париетальных) корковых нейронных систем разных полушарий (Andersen P., Anderson S.A., 1968; Ливанов М.Н., 1972; Lopes da Silva F.H. et al., 1973; Дубикайтис В.В., 1975; Свидерская Н.Е., 1987; Зенков Л.Р., Ронкин М.А., 1991; и др.).
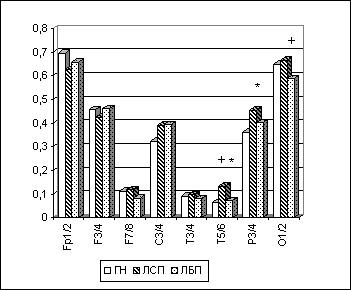
Рис. 1. Альфа-МКОГ при ОГ у лиц с парафилиями.
Примечания: Межгрупповые статистически достоверные различия (p<0.05 по LSD-тесту) в отдельных ЭЭГ-отведениях для пары "группа нормы-лица с парафилиями"(ГН-ЛСП) на рисунке обозначены звездочкой (*); для пары "группа нормы-лица без парафилий"(ГН-ЛБП) - кружком (о); для пары "лица с парафилиями- лица без парафилий" (ЛСП-ЛБП) - крестиком (+).
У лиц с парафилиями также имелся характерный (по отношению к ГН и ЛБП) зональный паттерн распределения ЛАСМ в коре головного мозга во всех ЭЭГ-диапазонах в различных функциональных состояниях, т.е. ЛАСМ у ЛСП был сравнительно меньше в лобно-центральных и больше в височных регионах неокортекса. Можно отметить, что данное повышение временной синхронизации ЭЭГ-ритмики в височных регионах хорошо согласуется с описанным выше увеличением в этой же области пространственной (межполушарной) синхронизации. Указанное снижение спектральной мощности в лобно-центральных областях неокортекса может быть обусловлено повышением десинхронизирующих (гиперактивационных) влияний, а повышение спектральной мощности в височных областях - усилением, напротив, гипоактивационных (синхронизирующих) влияний в этих регионах коры головного мозга (Beaumont J.G., 1983; Davidson R.J., 1988; и др.).
Кроме этого, ЛАСМ в тета-диапазоне при зрительно-пространственном задании у лиц с парафилиями достоверно (p<0.05) меньше в правом полушарии как по отношению к ГН в центральной и париетальной областях, так и относительно ЛБП в центральной и лобной областях (рис. 6.2, 6.3). В свою очередь, данный асимметрично повышенный уровень временной десинхронизации ЭЭГ-ритмики в тета-диапазоне в правом полушарии при зрительно-пространственном задании может быть интерпретирован (Beaumont J.G., 1983; Davidson R.J., 1988; De Toffol B. et al., 1990; Wheeler R.E. et al., 1993; и др.) как отражающий относительное повышение у лиц с парафилиями активационных влияний в лобно-центральных областях правого полушария.
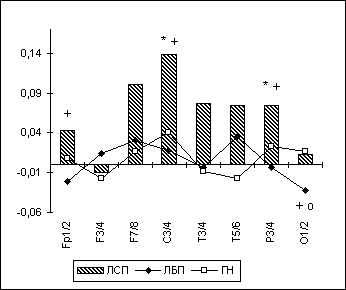
Рис. 2. Полушарная асимметрия по тета-ЛАСМ при ЗПЗ у лиц с парафилиями.
Примечания: На рисунке представлены значения индекса полушарной асимметрии (по ЛАСМ) в отдельных парах симметричных ЭЭГ-отведений ("ЛАСМ слева" минус "ЛАСМ справа") , т.е. положительные значения по оси ординат отражают относительно меньшие значения ЛАСМ справа и/или относительно большие значения ЛАСМ слева в симметричных ЭЭГ-отведениях, и наоборот. Межгрупповые статистически достоверные различия обозначены так же, как на рис. 6.1.
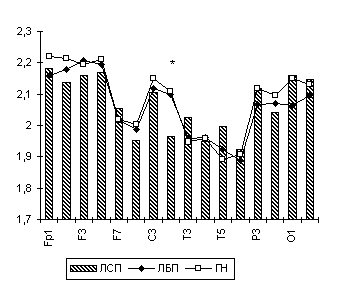
Рис. 3. Тета-ЛАСМ при ЗПЗ у лиц с парафилиями.
Примечания: Межгрупповые статистически достоверные различия обозначены так же, как на рис. 1.
Было проведено сопоставление изменений в ЭЭГ у исследуемых праворуких лиц с аномальным сексуальным поведением с нейропсихологическими синдромами, встречающимися у данной категории испытуемых. Для этого были выделены и сопоставлены между собой и с группой нормы (49 чел.) по ЭЭГ-характеристикам две клинические группы: лица с нейропсихологическим синдромом, отражающим нарушение когнитивных функций правого полушария (11 чел.), и лица с синдромом, отражающим нарушение когнитивных функций и левого, и правого полушарий с нарушением межполушарных взаимосвязей (15 чел.). Было выявлено, что только для первой ("правополушарной") группы был характерен паттерн, наблюдаемый у лиц с парафилиями, т.е. достоверное повышение межполушарной когерентности в задневисочных областях и снижение спектральной мощности ЭЭГ-ритмики в правом полушарии. Что, таким образом, подтверждает преимущественно правополушарную локализацию дисфункциональных ЭЭГ-сдвигов в головном мозге при парафилиях как в отношении спектральной мощности, так и в отношении межполушарной когерентности (и в том числе дает основание предполагать, что в изменении корково-подкорковых отношений в височной области наибольшее значение играют корковые правополушарные нейрональные механизмы).
Полученные результаты нейрофизиологического исследования вполне соотносятся с предположениями, сделанными по результатами клинического обследования данных больных, т.е. с предположениями о наличии у лиц с парафилиями органических процессов, связанных с нарушением деятельности структур височной доли и дисфункцией субдоминантного (правого) полушария. Учитывая, что основным отличием клинической картины у данной группы лиц с парафилиями (по сравнению с испытуемыми с аномальным сексуальным поведением без парафилий) было наличие практически у всех из них раннего или сочетанного органического поражения головного мозга, имеющего резидуальный характер, можно с большой долей уверенности предположить, что оно в основном и обусловило особенности наблюдаемого паттерна ЭЭГ.
Однако вопрос о том, связаны ли эти сдвиги в ЭЭГ с любыми или только с какими-либо особыми (в смысле нарушения нейрофункциональной организации мозга) вариантами резидуально-органического поражения, нуждался в прояснении. Это было сделано в работе, посвященной анализу особенностей ЭЭГ у лиц с резидуально-органическими поражениями головного мозга без аномального сексуального поведения.
2.2. Нейрофизиологические паттерны полушарных и корково-подкорковых отношений у лиц с резидуально-органическим поражением головного мозга
По методике, описанной выше, было проведено компьютерное ЭЭГ-обследование 40 лиц мужского пола с экзогенно-органическими поражениями головного мозга, находившихся на судебно-психиатрической экспертизе в ГНЦ СиСП им. В.П. Сербского 1, и группы нормы (ГН) в количестве 40 неврологически и психически здоровых праворуких мужчин. Были выделены две клинические группы лиц - с резидуальным (группа ЛРО, 11 чел.) и текущим (группа ЛТО, 29 больных) вариантами экзогенно-органического поражения головного мозга. У ЛРО отмечалось преобладание эмоционально-волевых расстройств в рамках церебрастенического и психопатоподобного синдромов, невротических и аффективных расстройств.
Полученные результаты указывали на то, что для ЛРО можно считать характерным значимое повышение в лобно-височных зонах межполушарной когерентности всех ЭЭГ-диапазонов, при снижении ее в затылочных областях (рис. 6.4), что, как указывалось выше, могло бы быть связано с изменением у них, как и у лиц с парафилиями, в этих регионах корково-подкорковых отношений. Кроме этого, у группы ЛРО отмечалось повышение преимущественно в лобных, центральных и височных областях правого полушария спектральной мощности высокочастотной (бета-1) и медленноволновой (дельта- и тета-) ЭЭГ-ритмики (Рис. 6.5). Что, в свою очередь, может отражать инактивационные изменения в правом полушарии.
1 Данное исследование было выполнено совместно со старшим научным сотрудником отделения экзогенно-органических расстройств Центра канд. мед. наук М.В.Усюкиной.
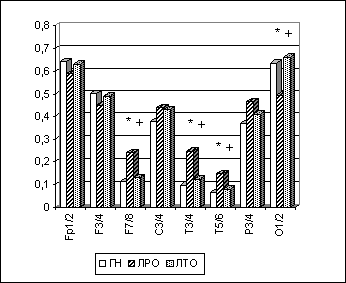
Рис. 4. Альфа-МКОГ при ОГ у лиц с органическим поражением головного мозга.
Примечания: Межгрупповые статистически достоверные различия (p<0.05 по LSD-тесту) в отдельных ЭЭГ-отведениях для пары "группа нормы-лица с резидуальным органическим поражением головного мозга"(ГН-ЛРО) на рисунке обозначены звездочкой (*); для пары "группа нормы-лица с текущим органическим поражением головного мозга"(ГН-ЛТО) - кружком (о); для пары "лица с резидуальным- лица с текущим органическим поражением головного мозга" (ЛРО-ЛТО) - крестиком (+).
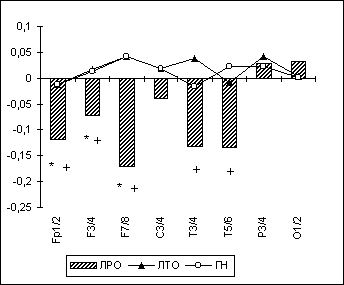
Рис. 5. Полушарная асимметрия по бета-ЛАСМ при ОГ у лиц с органическим поражением головного мозга.
Примечания: На рисунке представлены значения индекса полушарной асимметрии (по ЛАСМ) в отдельных парах симметричных ЭЭГ-отведений ("ЛАСМ слева" минус "ЛАСМ справа"). Межгрупповые статистически достоверные различия обозначены так же, как на рис. 4.
Учитывая описанные в первой части главы сведения и концепции об относительной специализации правополушарных лимбико-кортикальных систем на оценке эмоционально-отрицательных сенсорных образов, можно предположить у данных лиц с резидуальной органикой и асоциальным поведением определенную функциональную недостаточность правополушарных систем реагирования на отрицательные эмоциональные стимулы и ситуации. Подобная селективная дефицитарность была выявлена ранее в психофизиологических исследованиях (Patrick C.J., 1993) и у асоциальных психопатов, у которых она была связана с выраженностью одного из двух основных факторов асоциальной психопатии (Patrick С.J. et al. 1993), по критериям опросника "PCL-R" ("Psychopathy Checklist - Revised", 1991) - фактора "эмоциональной невовлеченности" ("emotional detachment").
Таким образом, очевидно, что если для резидуально-органических поражений головного мозга у лиц с асоциальным поведением без парафилий характерна гипоактивация правого полушария (и вероятная дефицитарность систем оценки эмоционально-отрицательных образов), то резидуально-органические поражения, сопровождаемые расстройствами сексуального влечения, являются, напротив, "правополушарно-гиперактивационными" по своим нейрофизиологическим механизмам (и, возможно, связаны уже с гиперактивацией систем оценки аверсивных стимулов). Последнее подтверждается и данными клинического анализа рассматриваемых лиц с парафилиями, у которых достоверно чаще встречались субдепрессивные и дистимические расстройства и дисфории.
2.3. Связь выявленных нейрофизиологических паттернов с дизонтогенетическими нарушениями в деятельности корковых и подкорковых структур головного мозга у лиц с парафилиями
Как установлено при ЭЭГ-исследованиях лиц с задержкой соматосексуального развития (Kaiser J., Gruzelier J., 1996), у них отмечается достоверно повышенный уровень когерентности ЭЭГ, что объясняется сниженным содержанием синапсов в коре вследствие более продолжительной их элиминации в процессе полового созревания (Huttenlocher P.R. et al., 1982; Saugstad L., 1994). Кроме этого, показано, что у мужчин с задержкой пубертата значимо более выражен шизотипический синдром "Избегания" ("Withdrawn"), который, в свою очередь, тесно связан с относительно большей активацией правого полушария (Gruzelier, J.H., Kaiser, J., 1996). Таким образом, есть определенные основания связывать формирование наблюдаемого у лиц с парафилиями паттерна ЭЭГ (т.е. повышение межполушарной когерентности и правополушарной гиперактивации) с дизонтогенетическими механизмами задержки у них соматосексуального развития, которые, в свою очередь, этиологически могут быть обусловлены ранним резидуальным органическим поражением головного мозга.
В пользу предположения о связи наблюдаемого нейрофизиологического паттерна с нарушением сексуального онтогенеза у мужчин говорит и то, что, как установлено, доминирование "правополушарных" функций характерно и для феминизированных мужчин (Кимура Д., 1992; Gruzelier J.H., 1994; и др.). Интересно, что у маскулинизированных женщин, согласно данным тех же исследований, наблюдается, наоборот, левополушарное доминирование, в то время как у "нормальных" мужчин и женщин существенной полушарной асимметрии не отмечается. Это может быть объяснено тем, что в норме развитие полушарий, по всей видимости, носит гармоничный и сбалансированный характер, а возникновение выраженной полушарной асимметрии может являться следствием гормонально-зависимых отклонений в развитии мозговых полушарных структур на ранних стадиях онтогенеза. Причем, возможно, что вследствие половых различий в нейроэндокринной регуляции одни и те же дизонтогенетические факторы (приводящие к расстройству в формировании и соответствующих мужских или женских половых признаков и характеристик) могут, вероятно, по-разному влиять на полушария головного мозга у мужчин и женщин. Так, у мужчин эти факторы могут вызывать торможение роста нейрональных структур левого полушария и компенсаторно большее развитие правого полушария ("тестостероновая гипотеза" - Geschwind N., Galaburda A.M., 1985), а у женщин, напротив, - большее торможение роста правого полушария и развитие "левополушарной" доминантности.
Учитывая это, можно было предположить, что у мужчин с сексуальными девиациями более выраженные расстройства половой идентичности сочетаются с более грубыми нарушениями корково-подкорковых и полушарных отношений.
Для проверки данного предположения было проведено следующее исследование. Праворукие испытуемые мужского пола с парафилиями, у которых была записана ЭЭГ, на основании результатов сексологического обследования и шкалирования были разделены на две группы. Первую клиническую группу составили 17 лиц без выраженных искажений половой идентичности (ЛБВИ). В эту группу были включены испытуемые, набравшие до 19 баллов по шкале оценки половой идентичности, которая была разработана в лаборатории судебной сексологии Центра. Во вторую группу вошли 13 лиц с выраженными искажениями половой идентичности (ЛВИ). Клинические группы были сопоставимы по возрасту и соотношению разных форм парафилий. Статистическая обработка параметров спектрально-когерентного анализа ЭЭГ (ЛАСМ, МКОГ) проводилась по описанной ранее схеме (ANOVA, LSD-тест). Клинические группы сравнивались между собой и с 49 праворукими лицами мужского пола из группы нормы (ГН).
Было обнаружено, что у ЛБВИ, по сравнению с ГН, достоверное повышение межполушарной когерентности в задневисочных и париетальных областях неокортекса отмечалось в тета- и альфа-диапазонах ЭЭГ при активном бодрствовании с открытыми глазами. У ЛВИ, по сравнению с ГН, статистически значимое повышение МКОГ в височных и париетальных зонах коры головного мозга количественно было более выражено и наблюдалось во всех ЭЭГ-диапазонах не только при ОГ, но и при других функциональных состояниях (рис. 6.6).
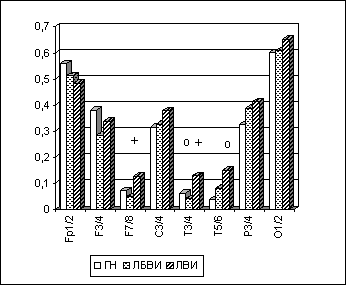
Рис. 6. Бета-МКОГ у лиц с искажениями половой идентичности.
Примечания: Межгрупповые статистически достоверные различия (p<0.05 по LSD-тесту) в отдельных ЭЭГ-отведениях для пары "группа нормы-лица с выраженными искажениями половой идентичности" (ГН-ЛВИ) на рисунке обозначены кружком (о); для пары "лица без выраженных искажений половой идентичности - лица с выраженными искажениями" (ЛБВИ-ЛВИ) - крестиком (+).
При анализе сдвигов в полушарной асимметрии (по ЛАСМ) было обнаружено, что клиническая группа ЛБВИ достоверно не отличалась от ГН, в то время как у лиц с выраженными нарушениями половой идентичности картина была совершенно другой. Отмечалось статистически значимое генерализованное (сочетание факторов "группа * полушарие") изменение полушарных отношений в группе ЛВИ во всех ЭЭГ-диапазонах при различных функциональных состояниях (прежде всего при ЗГ и ОГ). Спектральная мощность ЭЭГ у ЛВИ по сравнению с ГН была ниже в правом и/или выше в левом полушарии. Причем, в бета-1-диапазоне ЛАСМ у ЛВИ был достоверно выше в левом полушарии не только относительно ГН, но и относительно ЛБВИ (рис. 6.7).
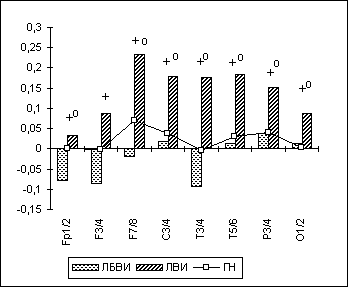
Рис. 7. Полушарная асимметрия по бета-ЛАСМ при ОГ у лиц с искажениями половой идентичности.
Примечания: На рисунке представлены значения индекса полушарной асимметрии (по ЛАСМ) в отдельных парах симметричных ЭЭГ-отведений ("ЛАСМ слева" минус "ЛАСМ справа"). Межгрупповые статистически достоверные различия обозначены так же, как на рис. 6.6.
Таким образом, исходя из полученных результатов, можно констатировать, что существенное изменение корково-подкорковых и полушарных отношений отмечается прежде всего у тех мужчин с парафилиями, у которых имеются выраженные нарушения половой идентичности. Этот факт, принимая во внимание тесную связь расстройств половой идентичности с психосексуальным дизонтогенезом, подтверждает значимость структурно-функциональных нарушений в мозге на ранних стадиях онтогенеза для формирования описанных нами паттернов патофизиологических сдвигов у лиц с сексуальными девиациями.
Раннее органическое поражение головного мозга, приводящее к формированию дизонтогенетических нарушений, может (при наличии определенной генетической предрасположенности) затрагивать не только корковые, но и подкорковые нейрональные структуры. Представляется, что в патофизиологических механизмах таких нарушений на подкорковом уровне могли бы иметь существенное значение дисфункциональные изменения нейрохимических регуляторных систем.
Как указывает А.Ф. Изнак (1997), скопления нейронов, продуцирующих нейротрансмиттеры, узко локализованы. Так, нервные клетки, синтезирующие серотонин, расположены в ядрах шва продолговатого мозга, норадренергические нейроны - в области голубого пятна, дофамин-продуцирующие клетки - в черной субстанции и мезолимбической области. Однако аксональные проекции этих нейронов широко распостранены в мозге. Это обусловливает возникающие при нарушении деятельности данных нейрональных систем изменения функционального состояния широкого круга мозговых структур. В контексте данной работы представляет особый интерес тот факт, что обильные серотонинергические проекции из дорзальных ядер шва получает, в частности, и стриатум (Molliver M.E., 1987; и др.). Одной же из основных функций стриатума, как известно, является организация сложных паттернов автоматизированных действий, нередко филогенетически древних по своему генезу (MacLean P.D., 1978; Хомская Е.Д., 1987; и др.), и могущих быть сопоставленными с поведенческими паттернами при обсессивных и компульсивных расстройствах.
Для объяснения механизмов связи аномального сексуального поведения с нарушениями в деятельности подкорковых структур и нейрохимическими дисфункциональными сдвигами необходимо рассмотреть особенности коморбидности парафилий. Так, установлена связь компульсивных вариантов девиантного сексуального поведения как с синдромом Туретта (Comings D.E., Comings D.G., 1982; Pitman R.K., 1983; Kerbeshian J. et al., 1985; Comings D.E., 1994), так и с обсессивно-компульсивными расстройствами (Coleman E., 1991; и др.), которые, в свою очередь, могут сочетаться и друг с другом (Pauls D.L., Leckman J.F., 1986; Leckman J.F. et al., 1992).
Согласно современным представлениям, синдром Туретта обусловлен прежде всего нарушением функций стриопаллидарной системы (Pauls D.L. et al., 1986; и др.) и генетически определенными изменениями метаболизма серотонина и дофамина (Comings D.E., 1990, 1994 и др.). В частности, обнаружено, что у больных с синдромом Туретта достоверно понижено содержание триптофана в крови (Leckman J.F. et al., 1984; Comings D.E., 1990), а также снижен уровень 5-HT в подкорковых регионах головного мозга (Anderson G.M. et al., 1992).
Патогенез обсессивно-компульсивных расстройств (ОКР) также во многом обусловлен нарушениями серотонинергических и норадренергических механизмов в мозге (МакГлинн Т., Меткалф Г.Л., 1989; Coleman E., 1991; Insel T.R., 1991; Hollander Е., 1993 и др.). На это указывают как данные о том, что у пациентов с ОКР в цереброспинальной жидкости выявлено повышение 5-HIAA (Insel T.R. et al., 1985), так и эффективность лечения больных с ОКР ингибиторами захвата серотонина (Insel T.R., 1991; и др.).
Как и при синдроме Туретта, патофизиологические механизмы обсессивно-компульсивных расстройств тесно связаны с нарушениями в деятельности базальных ганглиев. Существенное значение при этом, по всей видимости, имеют дисфункциональные изменения в системе "стриатум - лобные доли". Об этом говорит, в частности, уменьшение выраженности ОКР после хирургических операций, сопровождающихся пересечением проводящих путей от лобной коры к стриатуму (Bingley T., Persson A., 1978; Fodstad H. et al., 1982). Причем, как было выявлено в ПЭТ-исследованиях, при ОКР преобладают гиперактивационные изменения в хвостатом ядре и орбитальных областях лобной коры (Baxter L.R. et al., 1987, 1988; Nordahl T.E. et al., 1989; Swedo S.E. et al., 1989).
Таким образом, учитывая все эти факты, можно предположить, что общим патофизиологическим базисом для синдрома Туретта, обсессивно-компульсивных расстройств и компульсивных вариантов парафилий могли бы быть гиперактивационные дисфункциональные изменения в нейрональной системе "базальные ганглии - лобные доли", патогенез которых во многом обусловлен нарушениями серотонинергических и норадренергических регуляторных механизмов мозга.
Исходя из этого, относительное повышение активационных (десинхронизирующих ЭЭГ) влияний в лобно-центральных регионах головного мозга, обнаруженное нами у лиц с парафилиями, могло бы быть связано с наличием у них гиперактивационных изменений в системе "базальные ганглии - лобные доли", значимых для формирования компульсивных вариантов аномального сексуального поведения. Однако данное предположение нуждается в тщательной и всесторонней проверке.
2.4. Связь выявленных нейрофизиологических паттернов с особенностями конституционально обусловленных черт личности у испытуемых с парафилиями
Для изучения того, как выявленные изменения полушарных и корково-подкорковых отношений у лиц с парафилиями связаны с "базисными" (биологически обусловленными) характеристиками поведения и личности, было проведено следующее исследование. Праворуким испытуемым с аномальным сексуальным поведением (24 чел.) и лицам контрольной группы нормы (11 чел.) было предложено ответить на вопросы ряда соответствующих психологических тестов (Айзенка, Спилбергера, Цукермана, УСК, Русалова, Элерса и пр.). Только по двум из этих тестов были получены достоверные различия между клиническими группами лиц с аномальным сексуальным поведением с парафилиями (9 чел.) и без парафилий (15 чел.). Это "Опросник структуры темперамента" - ОСТ (Русалов В.М., 1989, 1990), количественно оценивающий предметно-ориентированые и коммуникативные характеристики темперамента, и опросник Элерса "Мотивация к достижению успеха - МДУ (разработан Ehlers T., 1964; адаптирован Котиком М.А., 1987), который оценивает силу мотивации к достижению цели, к успеху. Было выявлено, что у лиц с парафилиями (ЛСП) в сравнении с лицами без парафилий (ЛБП) и с нормой (ГН) достоверно снижены (LSD-test, p<0.05) как "эргичность" (ЭР), "пластичность" (П) и "темп" (Т) предметной деятельности по опроснику Русалова, так и "мотивация к достижению успеха" (МДУ) по опроснику Элерса (рис. 6.8). После этого в группе лиц с аномальным сексуальным поведение были вычислены коэффициенты корреляции (по Спирмену) между данными психометрическими показателями и параметрами МКОГ и полушарной асимметрии (по ЛАСМ). У лиц с аномальным сексуальным поведением снижение "мотивации к достижению успеха" достоверно (p<0.01) положительно коррелировало с относительно меньшими величинами тета-ЛАСМ в правом полушарии (и/или сравнительно большими величинами тета-ЛАСМ в левом) в центральных областях. В то же время снижение у них "эргичности" и "пластичности" предметной деятельности достоверно (p<0.05) коррелировало с повышением межполушарной когерентности в тета-, альфа- и бета-1-диапазонах ЭЭГ в височных и лобных областях головного мозга.
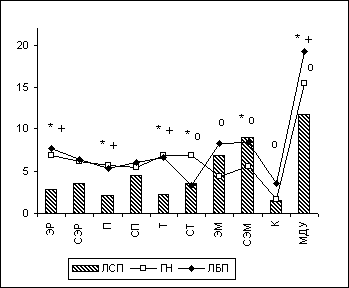
Рис. 8. Значения шкал опросников Русалова (ОСТ) и Элерса (МДУ) у лиц с парафилиями.
Примечания: По оси абсцисс представлены шкалы опросника Русалова (ОСТ): "эргичность" (ЭР), "социальная эргичность" (СЭР), "пластичность" (П), "социальная пластичность" (СП), "темп" (Т), "социальный темп" (СТ), "эмоциональность" (ЭМ), "социальная эмоциональность" (СЭМ), "контрольная шкала" (К). Кроме этого, представлена шкала опросника Элерса "мотивация к достижению успеха" (МДУ). Межгрупповые статистически достоверные различия обозначены так же, как на рис. 6.1.
Полученные результаты, по всей видимости, указывают на то, что характерное для лиц с парафилиями снижение "мотивации к достижению успеха" связано с изменением у них полушарной асимметрии в сторону большей активации правополушарных и/или меньшей активации левополушарных структур. То есть, как вытекает из этих данных и положений, рассмотренных в первой части этой главы, у лиц с расстройствами сексуального влечения, возможно, имеет место относительная функциональная недостаточность "левополушарной" лимбико-кортикальной системы, обеспечивающей мотивационные процессы организации "достижения" эмоционально-позитивного (и социально поощряемого), по сравнению с комплементарной ей "правополушарной" системой организации "избегания" эмоционально-отрицательного (в т.ч. и чувств "социальной" и сексуальной неудовлетворенности или неполноценности).
Кроме этого, можно предположить, что изменение корково-подкорковых отношений в височной области у лиц с парафилиями, приводящее к снижению степени относительной автономности деятельности височных корковых нейрональных систем разных полушарий и выражающееся в повышении МКОГ, обусловливает снижение у них "эргичности" и "пластичности" психических и поведенческих процессов. Это, в свою очередь, может лежать в основе характерной для лиц с расстройствами сексуального влечения устойчивой тенденции к появлению повторных, стереотипно воспроизводимых идеаторных образов и побуждений к совершению однотипных, автоматизированных действий. В пользу такого предположения говорят как данные В.М. Русалова, С.В. Калашникова (1988) о том, что со степенью пластичности, проявляющейся во всех сферах психики, отрицательно коррелирует лобно-затылочная когерентность ЭЭГ в правом полушарии, так и давно установленная связь ригидности психических процессов при эпилепсии преимущественно с нарушением деятельности корково-подкорковых структур височной доли (Williams D., 1969; Карлов В.А., 1990; и др.).
То есть у лиц с парафилиями, по-видимому, имеется, как указывал Г.Н Крыжановский и др. (1994), патофизиологический нейрональный базис для поэтапного формирования устойчивой "патологической системы". Однако, как вытекает из полученных результатов, нейрофизиологические механизмы функционирования такой системы могут быть связаны не столько с наличием спонтанно активирующегося "генератора патологически усиленного возбуждения" в головном мозге (Крыжановский Г.Н и др., 1994), сколько со снижением степени автономности деятельности височно-париетальных нейрональных систем разных полушарий и уменьшением относительной независимости протекания комплементарных по отношению друг к другу "лево-" и "правополушарных" нейропсихических процессов. Это, в свою очередь, может затруднять адекватное ситуационно-обусловленное "переключение" с "правополушарной" на "левополушарную" (и обратно) когнитивную, эмоциональную или мотивационную стратегию (т.е., снижать пластичность нейропсихических процессов и стереотипизировать поведение).
2.5. Сравнительный анализ нейрофизиологических паттернов полушарных, зональных и корково-подкорковых отношений у лиц с различными формами парафилий
Учитывая литературные данные о различии отдельных форм парафилий (эксгибиционизм, педофилия versus сексуальный садизм) по полушарной локализации латерализованных церебральных нарушений (Flor-Henry P., 1988, 1991; Langevin R. et al., 1988; Ether R. et al. 1996; и др.) нами на данной выборке праворуких лиц мужского пола с парафилиями был проведен сравнительный анализ особенностей ЭЭГ у испытуемых с разными формами парафилий: у лиц с садизмом - ЛСС (20 чел.) и у лиц с парафилиями без садизма - ЛБС (17 чел., в том числе 10 чел. с педофилией и 7 с эксгибиционизмом), и у праворуких мужчин из группы нормы - ГН (49 чел.).
У обеих групп ЛСС и ЛБС наблюдалось снижение МКОГ по отношению к ГН в лобных и центральных регионах. Однако только у лиц с садизмом по сравнению с ГН отмечалось выраженное достоверное (p<0.05) повышение межполушарной когерентности в височных областях неокортекса в тета-, альфа- и бета-1-диапазонах ЭЭГ при различных функциональных состояниях (рис. 6.9). У лиц же с парафилиями без садистического компонента о повышении МКОГ можно говорить в основном как о тенденции (МКОГ достоверно выше относительно ГН в височных областях только в бета-диапазоне при открытых глазах).
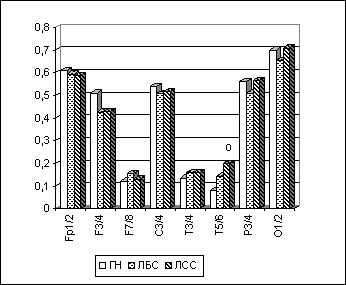
Рис. 9. Тета-МКОГ при ОГ у лиц с различными формами парафилий.
Примечания: Межгрупповые статистически достоверные различия (p<0.05 по LSD-тесту) в отдельных ЭЭГ-отведениях для пары "группа нормы-лица с садизмом"(ГН-ЛСС) обозначены кружком (о).
У ЛСС (по сравнению с ГН) отмечается и определенный зонально-зависимый паттерн распределения спектральной мощности в коре головного мозга во всех ЭЭГ-диапазонах в различных функциональных состояниях, т.е. ЛАСМ у лиц с садизмом сравнительно меньше в лобно-центральных и больше в височных регионах неокортекса. У ЛБС по отношению к ГН также наблюдается сходный зонально-зависимый паттерн распределения ЛАСМ, но только в дельта- и бета-диапазонах.
У лиц с разными формами расстройств сексуального влечения наблюдается сходный по типу, но различный по заинтересованности полушарий паттерн измененных полушарных отношений. У лиц с садизмом отмечается относительно большая временная десинхронизация ЭЭГ-ритмов в центральной и лобной областях правого полушария не только в тета-диапазоне при зрительно-пространственном задании (как и в целом у лиц с парафилиями), но и в альфа- и бета-1-диапазонах в различных функциональных состояниях (рис. 6.10). Это может говорить о существенном повышении именно у садистов активационных влияний в этих зонах правого полушария. У лиц же с педофилией и эксгибиционизмом отмечается уже относительно большая временная синхронизация ЭЭГ-ритмики в бета-1-диапазоне в лобных и височных регионах левого полушария, что может быть связано с определенным снижением активационных влияний в соответствующих областях левого полушария.
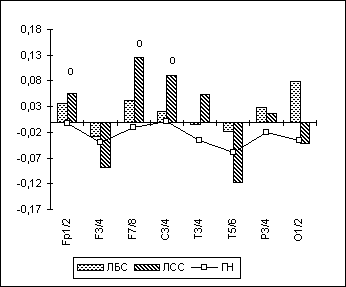
Рис. 10. Полушарная асимметрия по альфа-ЛАСМ при ЗГ у лиц с различными формами парафилий.
Примечания: На рисунке представлены значения индекса полушарной асимметрии (по ЛАСМ) в отдельных парах симметричных ЭЭГ-отведений ("ЛАСМ слева" минус "ЛАСМ справа"). Межгрупповые статистически достоверные различия обозначены так же, как на рис. 6.9.
Кроме этого, у лиц с садизмом по отношению к норме отмечаются достоверно большие величины правополушарной ЭЭГ-реакции, т.е. более выраженное гиперактивационное снижение спектральной мощности в альфа-диапазоне в правом полушарии (особенно в затылочной области) при счете в уме. (рис. 6.11). Возможно, это связано с тем, что лица с садизмом используют правополушарную когнитивную стратегию и в ситуациях, предполагающих в норме преимущественную активацию левого полушария (при счете в уме). Не исключено, что эта закономерность, касающаяся преобладания у садистов "правополушарных" процессов как в адекватных, так и в неадекватных этому ситуациях, распространяется не только на когнитивную, но и на эмоционально-мотивационную сферу психики.
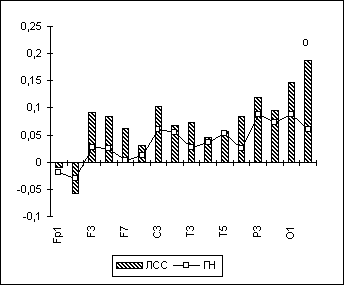
Рис. 11. ЭЭГ-реакция на СЧЕТ по альфа-ЛАСМ у сексуальных садистов.
Примечания: На рисунке представлены значения индекса ЭЭГ-реакции (по ЛАСМ) на арифметический счет в уме с закрытыми глазами ("ЛАСМ при ЗГ" минус "ЛАСМ при СЧЕТЕ") т.е., положительные значения по оси ординат отражают уменьшение ЛАСМ при СЧЕТЕ по сравнению с ЗГ. Межгрупповые статистически достоверные различия обозначены так же, как на рис. 6.9.
Полученные данные хорошо согласуются с имеющимися в литературе результатами исследований лиц с несадистическими формами парафилий (педофилия, эксгибиционизм), у которых, согласно данным P. Flor-Henry и сотр. (1988, 1991), обнаруживаются ЭЭГ-изменения, связанные с левополушарной дисфункцией со снижением активации в лобно-височных регионах. Однако тот ЭЭГ-паттерн, который мы впервые описали у лиц с садизмом, свидетельствует о том, что у них аналогичная по типу полушарная асимметрия связана с существенным повышением активации в правом полушарии. Это, с одной стороны, вполне соотносится с результатами компьютерной и магниторезонансной томографии, согласно которым у садистов выявляются существенные структурные изменения именно в правом полушарии (Langevin R. et al., 1988; Hucker S.J. et al., 1988; Ether R. et al, 1996). С другой стороны, это указывает на возможную гиперактивацию у садистов правополушарных лимбико-кортикальных систем, обеспечивающих отрицательное эмоциональное реагирование.
Кроме того, как уже обсуждалось, гиперактивация правого полушария у садистов может при определенных условиях на уровне "базисных" мотивационных процессов способствовать превалированию стремления "избежать" или "избавиться" от эмоционально-отрицательных состояний (в том числе чувства сексуальной неудовлетворенности и "социальной" фрустрации), а на когнитивном уровне - к созданию "особых", "индивидуальных" (и даже "аномальных" и "асоциальных") моделей поведенческой активности. Все это могло бы иметь существенное значение в механизмах формирования у садистов аномального агрессивного сексуального поведения путем. В частности, путем создания патофизиологических предпосылок, которые облегчают развитие устойчивого доминирования в сексуальном поведении таких индивидуумов аномальных (асоциальных) способов избавления от остро переживаемых ими чувств сексуальной неудовлетворенности и социальной ущербности.
3. Концептуальная модель психо- и нейрофизиологических механизмов аномального сексуального поведения
На основе анализа литературных и собственных данных можно предложить следующую концепцию нейрофизиологических механизмов аномального сексуального поведения.
Согласно ей, раннее резидуальное органическое поражение головного мозга сопровождается структурно-функциональными нарушениями в подкорковых и корковых нейрональных структурах мозга. Некоторые варианты этих дисфункциональных сдвигов могут являться церебральными (патофизиологическими) факторами предиспозиции к возникновению расстройств сексуального влечения. Кроме церебральных факторов, для формирования аномального сексуального поведения необходимы также определенные патопсихологические условия. У каждого отдельного индивидуума с девиантным сексуальным поведением сочетание и соотношение патопсихологических и патофизиологических предиспозиционных факторов является индивидуальным и определяется конкретными особенностями онтогенеза и условиями жизнедеятельности.
Морфофункциональные нарушения в подкорковых структурах при рассматриваемых вариантах раннего органического поражения головного мозга могут касаться как гипоталамуса и миндалевидного комплекса, так и базальных ганглиев. Причем патофизиологические сдвиги в деятельности данных подкорковых образований, по всей видимости, имеют как постоянные (тонические), так и пароксизмальные (фазические) составляющие.
Дисфункциональные сдвиги в гипоталамусе могут приводить как к общим изменениям сексуальности в виде гипер- или гипосексуальности, так и к девиациям в способах и формах сексуальной активности, характерных для особей мужского или женского пола (что, вероятно, имеет особое значение в генезе гомосексуальности).
Нарушение деятельности структур миндалевидного комплекса способно существенно изменять как мотивационный анализ и эмоциональную оценку сенсорных сексуально-ассоциированных стимулов, так и инициацию адекватных паттернов сексуальной активности в зависимости от наличной ситуации и конкурирующих мотиваций. Поэтому дисфункциональные сдвиги в структурах миндалевидного комплекса могут как создавать патофизиологические предпосылки для формирования (в том числе и по механизму импринтинга) атипичных сексуальных предпочтений (к детям, животным, неодушевленным предметам и пр.), так и облегчать инициацию сексуальной активности в неподходящей для этого ситуации (в общественных местах и т.п.) или в социально-неприемлемых и аномальных формах (с использованием насилия, ненормативных способов сексуального удовлетворения и пр.).
Дисфункция структур базальных ганглиев (и связанных с ними нейрональных систем лобных долей) может создавать условия для реализации сексуальной активности в виде автоматизированных навязчивых действий, т.е. способствовать формированию компульсивных форм аномального сексуального поведения. Выявленная в наших исследованиях относительная гиперактивация лобно-центральных регионов головного мозга у лиц с парафилиями может свидетельствовать о преобладании при компульсивных формах парафилий гиперактивационных вариантов дисфункциональных изменений в системе "базальные ганглии-лобные доли". При этом данные дисфункциональные сдвиги могут быть одним из следствий патологических изменений в церебральных нейрохимических (серотонинергической, норадренергической и пр.) системах, регулирующих деятельность этих подкорковых и корковых структур мозга. Патогенез нейрохимических сдвигов, по всей видимости, обусловлен сочетанием определенных вариантов раннего органического поражения головного мозга с индивидуальной генетической предрасположенностью к возникновению изменений на нейрохимическом уровне.
У человека поведение, направленное на удовлетворение тех или иных биологических (в том числе и сексуальных) потребностей, невозможно без участия высших регуляторных корковых механизмов. Поэтому представляется, что для реализации в поведении и репрезентации в сознании тех или иных аномальных способов и форм сексуальной активности (патофизиологический базис для формирования которых может определяться дисфункцией рассмотренных подкорковых структур) необходимо и нарушение деятельности корковых нейрональных систем.
При этом, можно предположить, что такие дизонтогенетически обусловленные патофизиологические сдвиги на корковом уровне регуляции поведения могут формироваться в основном у мужчин, а не у женщин, что могло бы объяснить встречаемость расстройств сексуального влечения преимущественно у лиц мужского пола. Это, вероятно, связано с тем, что, как обсуждалось нами ранее, у мужчин дизонтогенетически уязвимы прежде всего "мотивационные" лобные лимбико-кортикальные системы регуляции поведения, в то время как у женщин уязвимы для дизонтогенетических факторов прежде всего лимбико-кортикальные системы, ответственные за анализ и оценку эмоционально-ассоциированных сенсорных стимулов (что, видимо, и обусловливает существенно более частую встречаемость у женщин аффективных расстройств). Кроме этого, как следует из анализа литературных данных, у мужчин и женщин, вследствие связанных с полом различий в нейроэндокринной регуляции, одни и те же дизонтогенетические факторы могут приводить к существенно различающимся (и даже противоположным) изменениям в паттернах полушарных отношений.
Механизмы формирования нейрофизиологических сдвигов в корковых структурах у мужчин могут быть представлены следующим образом. Определенные варианты органического поражения головного мозга на ранних стадиях онтогенеза способны приводить к задержке соматосексуального развития индивидуума. В свою очередь, задержка соматосексуального развития, во-первых, может вызывать снижение количества синапсов в коре головного мозга вследствие более продолжительной их элиминации в процессе полового созревания и соответствующее изменение корково-подкорковых взаимоотношений с повышением когерентности ЭЭГ. И, во-вторых, задержка пубертата у мужчин способна приводить к замедлению роста левого полушария и компенсаторно большему развитию и относительной гиперактивации структур правого полушария со сравнительно большей десинхронизацией ЭЭГ в правом полушарии. Причем общий дизонтогенетический базис может обуславливать у мужчин с задержкой пубертата сочетание описанного нейрофизиологического паттерна с феминизацией психических и соматических характеристик.
Результаты наших исследований не только укладываются в данную общую концептуальную схему, но и позволяют применить и конкретизировать эти положения в отношении мужчин с расстройствами сексуального влечения. Так, нарушение корково-подкорковых отношений и соответствующее повышение межполушарной когерентности ЭЭГ у лиц с парафилиями характерно прежде всего для задневисочных областей головного мозга. Изменение же полушарных отношений у мужчин с расстройствами сексуального влечения происходит как за счет повышения активационных влияний в лобно-центральных областях правого полушария (у садистов), так и за счет относительного снижения активации в этих регионах в левом полушарии (у лиц с педофилией или эксгибиционизмом). Наиболее существенные сдвиги в корково-подкорковых и полушарных отношениях отмечаются у тех мужчин с парафилиями, у которых имеются выраженные дизонтогенетические нарушения, проявляющиеся, в частности, расстройствами половой идентичности.
Анализ собственных и литературных данных позволяет предположить, что выявленные нами изменения в корковых полушарных системах могут определять некоторые особенности "базисных" (биологически обусловленных) черт личности и характеристик протекания мотивационных, эмоциональных и когнитивных процессов.
Уменьшение относительной автономности деятельности корковых нейрональных систем разных полушарий в височной области, возможно, создает патофизиологический базис для снижения пластичности нейропсихических процессов у лиц с парафилиями. Сочетаемая с этим дисфункциональная гиперактивация правого полушария (и/или гипоактивация левого) может обусловливать определенные устойчивые сдвиги в когнитивной, эмоциональной и мотивационной сферах психики.
Так, у лиц с парафилиями в когнитивной сфере может формироваться предрасположенность к созданию стереотипных "аномальных" схем сексуального поведения; в эмоциональной сфере - повышенная и гипопластичная эмоционально-отрицательная реактивность; в мотивационной сфере - устойчивое снижение мотивации к достижению позитивных социально одобряемых целей и/или повышение стремления к избавлению от неприятных и фрустрирующих индивидуума стимулов и ситуаций.
Как вытекает из полученных нами результатов, описанные патофизиологические сдвиги на корковом уровне имеют особенно важное значение в патогенезе агрессивного садистического поведения. Эти изменения способствуют, как предполагается, формированию, устойчивому доминированию и реализации в поведении таких индивидуумов ряда аномальных (асоциальных) потребностей. В частности, потребности избавиться от чувства сексуальной неудовлетворенности и социальной ущербности путем совершения стереотипных насильственных агрессивных действий.
В целом, можно констатировать, что церебральным фактором предиспозиции к формированию аномального сексуального поведения, по всей видимости, является дизонтогенетически обусловленный комплекс определенных патофизиологических сдвигов как на подкорковых, так и на корковых уровнях регуляции поведения. Особенности, взаимосвязь и соотношение таких дисфункциональных нарушений различны при разных формах расстройств сексуального влечения и у разных индивидуумов и определяются как структурно-функциональными характеристиками раннего резидуального поражения головного мозга, так и конкретными условиями онтогенеза.